quarta-feira, 25 de novembro de 2015
Mutação Gênica
Todos os dias as suas células produzem proteínas que contêm aminoácidos em uma certa seqüência. Imagine, por exemplo, que em um certo dia uma célula da epiderme de sua pele produza uma proteína diferente. Suponha também que essa proteína seja uma enzima que atue em uma reação química que leva a produção de um pigmento amarelo em vez do pigmento normalmente encontrado na pele, a melanina. Essa célula se multiplica e de repente aparece uma mancha amarelada em sua pele. Provavelmente essa proteína poderá ter sofrido uma alteração em sua seqüência de aminoácidos, tendo havido a substituição de um aminoácido por outro, o que acarretou uma mudança em seu mecanismo de atuação e, como conseqüência levou à produção de um pigmento de cor diferente. Agora, como a seqüência de aminoácidos em uma proteína é determinada pela ação de um certo gene que conduz à síntese do pigmento.
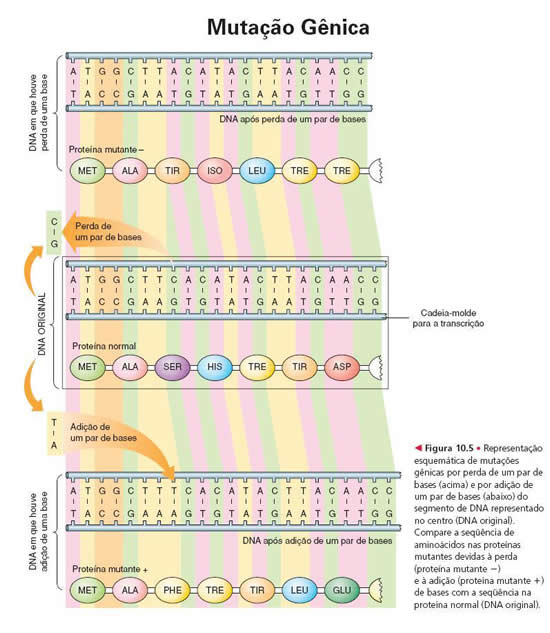
Essa alteração na seqüência de bases na molécula de DNA constituinte do gene é que se chama de mutação gênica.


O albinismo é causada por uma mutação na enzima tirozinase que transforma o aminoácido tirozina em pigmento da pele, a melanina. Esta doença ocorre em animais e nas plantas e é hereditária.
A mutação e suas conseqüências
Se a alteração na seqüência de aminoácidos na proteína não afetar o funcionamento da molécula e não prejudicar o organismo, de modo geral ela passa despercebida, é indiferente.
Outras vezes, a alteração leva a um favorecimento. Imagine, por exemplo, que uma certa célula do seu intestino passe a produzir uma enzima chamada celulase, capaz de digerir a celulose dos vegetais que você come. provavelmente a mutação que levou a esse erro será vantajosa para você, que poderá eventualmente até alimentar-se de papel picado.
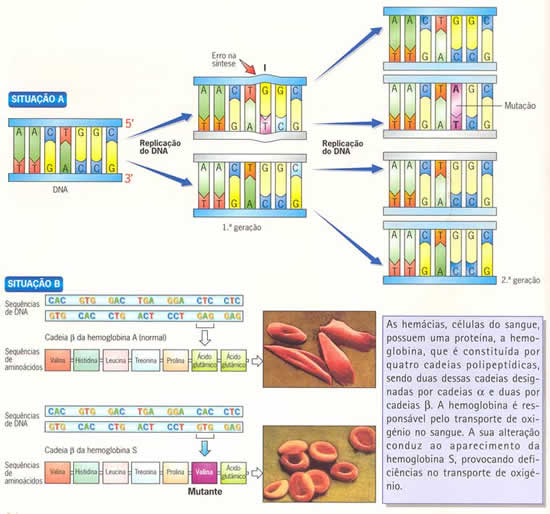
Muitas vezes, porém, a mutação pode ser prejudicial. Na anemia falciforme, a substituição do aminoácido ácido glutâmico pelo aminoácido valina, em uma das cadeias de hemoglobina, conduza a uma alteração na forma da proteína toda. Essa alteração muda o formato do glóbulo vermelho, que passa a ser incapaz de transportar oxigênio. Outra conseqüência, grave, é que hemácias com formato de foice grudam umas nas outras nos capilares sangüíneos, o que pode provocar obstruções no trajeto para os tecidos.
As mutações são hereditárias
Dependendo da célula em que a mutação ocorre, ela pode ser transmitida à descendência. Nas suposições que fizemos, relacionadas ao pigmento da pele e à enzima celulase, evidentemente que não ocorrerá a transmissão dos genes mutantes para os filhos.
Trata-se de mutações somáticas, ou seja, ocorreram em células não envolvidas na confecção de gametas.
Já a mutação que conduziu à anemia falciforme, deve ter ocorrido, no passado, em células da linhagem germinativa de algum antepassado. O gene anômalo, então sugerido, deve ter sido transportado por um gameta e daí se espalhou pela espécie humana.
As causas das mutações
De maneira geral, as mutações ocorrem como conseqüência de erro no processo de duplicação do DNA. Acontecem em uma baixíssima freqüência. Muitas delas, inclusive, são corrigidas por mecanismos especiais, como, por exemplo, a ação do gene p53 que evita a formação de tumores.
Há, no entanto, certos agentes do ambiente que podem aumentar a taxa de ocorrência de erros genéticos. Entre esses agentes mutagênicos podemos citar: substâncias existentes no fumo, os raios X, a luz ultravioleta, o gás mostarda, ácido nitroso e algumas corantes existentes nos alimentos. Não é à toa que, em muitos países, é crescente a preocupação com a diminuição da espessura da camada do gás ozônio (O3), que circunda a atmosfera terrestre. Esse gás atua como filtro de luz ultravioleta proveniente do Sol. Com a diminuição da sua espessura, aumenta a incidência desse tipo de radiação, o que pode afetar a pele das pessoas. Ocorrem lesões no material genético, que podem levar a certos tipos de câncer de pele. http://www.sobiologia.com.br/conteudos/Citologia2/AcNucleico10.php
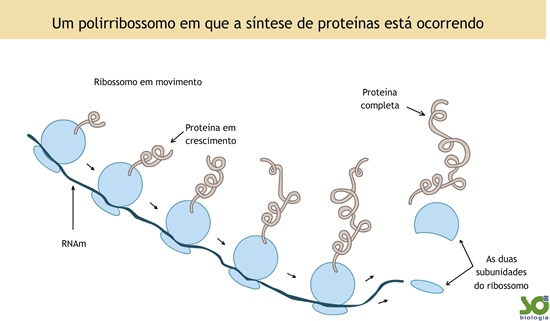
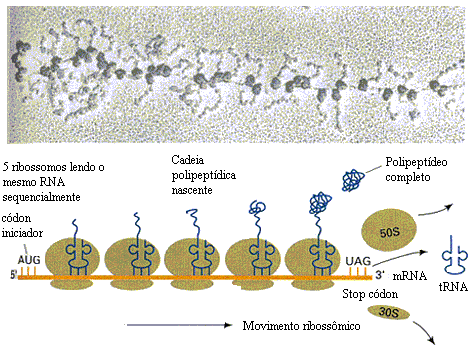
Os polirribossomos
Em algumas células, certas proteínas são produzidas em grande quantidade. Por exemplo, a observação de glândulas secretoras de certos hormônios de natureza protéica (que são liberados para o sangue, indo atuar em outros órgãos do mesmo organismo) mostra, em certos locais, uma fileira de ribossomos efetuando a leitura do mesmo RNA mensageiro. Assim, grandes quantidades da mesma proteína são produzidas.
Ao conjunto de ribossomos, atuando ao longo se um RNAm, dá-se o nome de polirribossomos.
Possíveis destinos das proteínas
O RNAm, ao sair do nucleo pode seguir dois destinos:
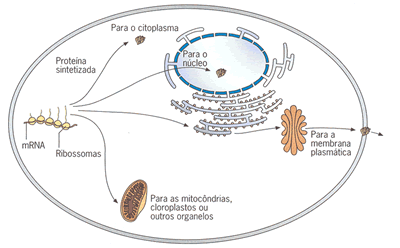
Síntese de proteínas no núcleo celular?
No núcleo de todas as células eucarióticas, a produção de RNA se da a partir do molde de uma fita do DNA. Então, o RNA sofre algumas alterações e segue para o citoplasma onde se associa aos ribossomos para a formação das proteínas. Acreditava-se que a síntese de proteínas (tradução) ocorresse somente no citoplasma, mas em recente trabalho publicado na prestigiada revista Science foi demonstrado que os elementos necessários à tradução se associam no núcleo, onde proteínas seriam formadas. Além disso os pesquisadores constataram que as estruturas responsáveis pela tradução estão em atividade no núcleo celular.http://www.sobiologia.com.br/conteudos/Citologia2/AcNucleico9.php
|
RNA - Tradução passo a passo
A tradução é um processo no qual haverá a leitura da mensagem contida na molécula de RNAm pelos ribosomo, decodificando a linguagem de ácido nucleico para a linguagem de proteína.
Cada RNAt em solução liga-se a um determinado aminoácido, formando-se uma molécula chamada aminoacil-RNAt, que conterá, na extremidade correspondente ao anticódon, um trio de códon do RNAm.
Para entendermos bem este processo, vamos admitir que ocorra a síntese de um peptídeo contendo apenas sete aminoácidos, o que se dará a partir da leitura de um RNAm contendo sete códons (21 bases hidrogenadas). A leitura (tradução) será efetuada por um ribossomo que se deslocará ao longo do RNAm.
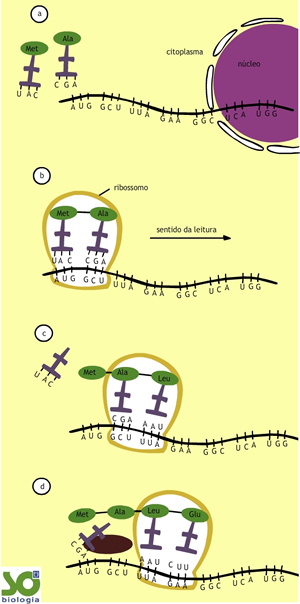
Esquematicamente na síntese proteica teríamos:
- Um RNAm, processado no núcleo, contendo sete códons (21 bases hidrogenadas) se dirige ao citoplasma.
- No citoplasma, um ribossomo se liga ao RNAm na extremidade correspondente ao início da leitura. Dois RNAt, carregando os seus respectivos aminoácidos (metionina e alanina), prendem-se ao ribossomo. Cada RNAt liga-se ao seu trio de bases (anticódon) ao trio de bases correspondentes ao códon do RNAm. Uma ligação peptídica une a metionina à alanina.
- O ribossomo se desloca ao longo do RNAm. O RNAt que carregava a metionina se desliga do ribossomo. O quarto RNAt, transportando o aminoácido leucina, une o seu anticódon ao códon correspondente do RNAm. Uma ligação peptídica é feita entre a leucina e a alanina.
- O ribossomo novamente se desloca. O RNAt que carregava a alanina se desliga do ribossomo. O quarto RNAt, transportando o aminoácido ácido glutâmico encaixa-se no ribossomo. Ocorre a união do anticódon desse RNAt com o códon correspondente do RNAm. Uma ligação peptídica une o ácido glutâmico à leucina.
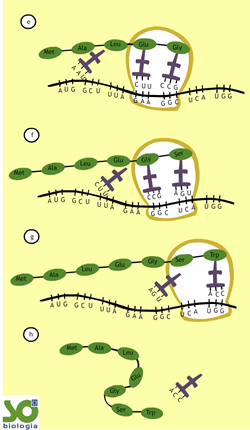
- Novo deslocamento do ribossomo. O quinto RNAt, carregando a aminoácido glicina, se encaixa no ribossomo. Ocorre a ligação peptídica da glicina com o ácido glutâmico.
- Continua o deslocamento do ribossomo ao longo do RNAm. O sexto RNAt, carregando o aminoácido serina, se encaixa no ribossomo. Uma liogação peptídica une a serina à glicina.
- Fim do deslocamento do ribossomo. O último transportador , carregando o aminoácido triptofano, encaixa-se no ribossomo. Ocorre a ligação peptídica do triptofano com a serina. O RNAt que carrega o triptofano se separa do ribossomo. O mesmo ocorre com o transportador que portava a serina.
- O peptídeo contendo sete aminoácidos fica livre no citoplasma. Claro que outro ribossomo pode se ligar ao RNAm, reiniciando o processo de tradução, que resultará em um novo peptídio. Perceba, assim, que o RNAm contendo sete códons (21 bases nitrogenadas) conduziu a síntese de um peptídeo formado por sete aminoácidos.http://www.sobiologia.com.br/conteudos/Citologia2/AcNucleico8.php
Tradução: Síntese de Proteínas
Tradução é o nome utilizado para designar o processo de síntese de proteínas. Ocorre no citoplasma com a participação, entre outros, de RNA e de aminoácidos.
Quem participa da síntese de proteínas?
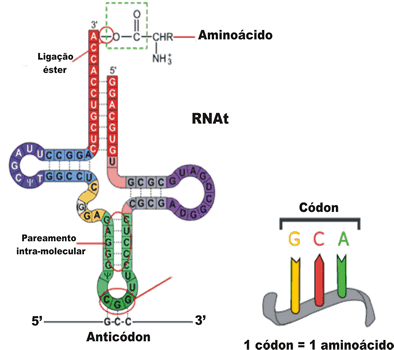
Cístron (gene) é o segmento de DNA que contém as informações para a síntese de um polipeptídeo ou proteína.
O RNA produzido que contém uma seqüência de bases nitrogenadas transcrita do DNA é um RNA mensageiro.
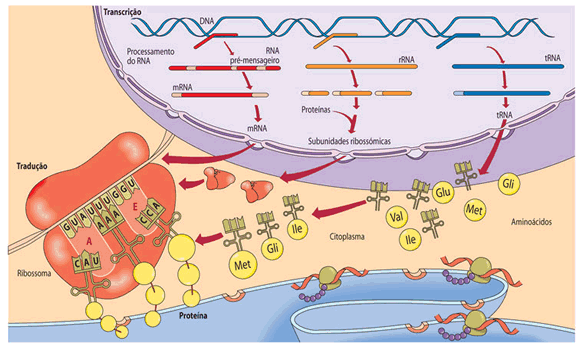
No citoplasma, ele será um dos componentes participantes da síntese de proteínas, juntamente com outros dois tipos de RNA, todos de fita simples e produzidos segundo o mesmo processo descrito para o RNA mensageiro:
RNA ribossômico, RNAr. Associando-se a proteínas, as fitas de RNAr formarão os ribossomos, orgânulos responsáveis pela leitura da mensagem contida no RNA mensageiro;
|
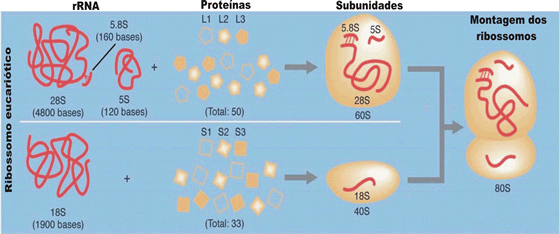
RNAs transportadores, RNAt. Assim chamados porque serão os responsáveis pelo transporte de aminoácidos até o local onde se dará a síntese de proteínas junto aos ribossomos. São moléculas de RNA de fita simples, de pequeno tamanho, contendo, cada uma, cerca de 75 a 85 nucleotídeos. Cada fita de RNAt torce-se sobre si mesma, adquirindo o aspecto visto na figura abaixo.
Duas regiões se destacam em cada transportador: uma é o local em que se ligará o aminoácido a ser transportado e a outra corresponde ao trio de bases complementares (chamado anticódon) do RNAt, que se encaixará no códon correspondente do RNAm.
Anticódon é o trio de bases do RNAt, complementar do códon do RNAm.
|
O código genético
A mensagem genética contida no DNA é formada por um alfabeto de quatro letras que correspondem aos quatro nucleotídeos: A, T, C e G. Com essas quatros letras é preciso formar “palavras” que possuem o significado de “aminoácidos”. Cada proteína corresponde a uma “frase” formada pelas “palavras”, que são os aminoácidos. De que maneira apenas quatro letras do alfabeto do DNA poderiam ser combinadas para corresponder a cada uma das vinte “palavras” representadas pelos vinte aminoácidos diferentes que ocorrem nos seres vivos.
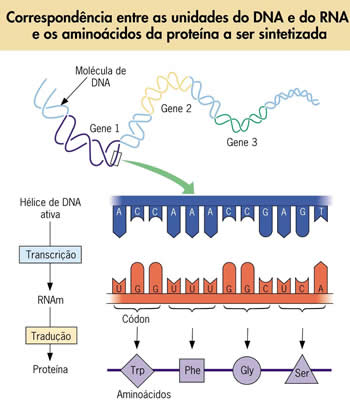
Uma proposta brilhante sugerida por vários pesquisadores, e depois confirmada por métodos experimentais, foi a de que cada três letras (uma trinca de bases) do DNA corresponderia uma “palavra”, isto é, um aminoácido. Nesse caso, haveria 64 combinações possíveis de três letras, o que seria mais do que suficiente para codificar os vinte tipos diferentes de aminoácidos (matematicamente, utilizando o método das combinações seriam, então, 4 letras combinadas 3 a 3, ou seja, 43 = 64 combinações possíveis).
O código genético do DNA se expressa por trincas de bases, que foram denominadas códons. Cada códon, formado por três letras, corresponde a um certo aminoácido.
A correspondência entre o trio de bases do DNA, o trio de bases do RNA e os aminoácidos por eles especificados constitui uma mensagem em código que passou a ser conhecida como “código genético”.
Mas, surge um problema. Como são vinte os diferentes aminoácidos, há mais códons do que tipos de aminoácidos! Deve-se concluir, então, que há aminoácidos que são especificados por mais de um códon, o que foi confirmado. A tabela abaixo, especifica os códons de RNAm que podem ser formados e os correspondentes aminoácidos que especificam.
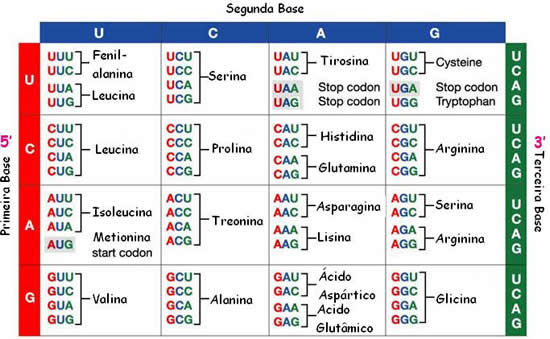
Dizemos que o código genético é universal, pois em todos os organismos da Terra atual ele funciona da mesma maneira, quer seja em bactérias, em uma cenoura ou no homem.
O códon AUG, que codifica para o aminoácido metionina, também significa início de leitura, ou seja, é um códon que indica aos ribossomos que é por esse trio de bases qe deve ser iniciada a leitura do RNAm.
Note que três códons não especificam nenhum aminoácido. São os códons UAA, UAG e UGA, chamados de códons e parada durante a “leitura” (ou stop códons) do RNA pelos ribossomos, na síntese protéica.
Diz-se que o código genético é degenerado porque cada “palavra” (entenda-se aminoácido) pode ser especificada por mais de uma trinca.http://www.sobiologia.com.br/conteudos/Citologia2/AcNucleico6.php
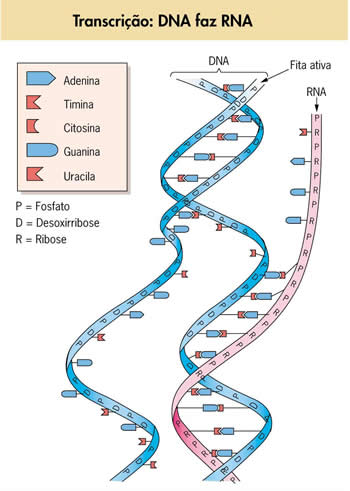
Transcrição da informação genética
A síntese de RNA (mensageiro, por exemplo) se inicia com a separação das duas fitas de DNA. Apenas uma das fitas do DNA serve de molde para a produção da molécula de RNAm. A outra fita não é transcrita. Essa é uma das diferenças entre a duplicação do DNA e a produção do RNA.
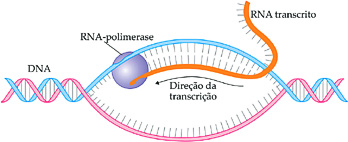
As outras diferenças são:
- os nucleotídeos utilizados possuem o açúcar ribose no lugar da desoxirribose;
- há a participação de nucleotídeos de uracila no lugar de nucleotídeos de timina. Assim, se na fita de DNA que está sendo transcrita aparecer adenina, encaminha-se para ela um nucleotídeo complementar contendo uracila;
Imaginando um segmento hipotético de um filamento de DNA com a seqüência de bases:
DNA- ATGCCGAAATTTGCG
O segmento de RNAm formado na transcrição terá a seqüência de bases:
RNA- UACGGCUUUAAACGC
Em uma célula eucariótica, o RNAm produzido destaca-se de seu molde e, após passar por um processamento, atravessa a carioteca e se dirige para o citoplasma, onde se dará a síntese protéica. Com o fim da transcrição, as duas fitas de DNA seu unem novamente, refazendo-se a dupla hélice.
Replicação, Transcrição e Tradução
Replicação, transcrição e tradução são processos que ocorrem com os ácidos nucléicos e que são essenciais para o funcionamento das nossas células. Em tópicos anteriores, já vimos a estrutura geral dosácidos nucléicos, a estrutura do DNA e sua organização em cromossomos e os diferentes tipos de RNA. Reveja todos esses tópicos para poder compreender melhor os tópicos em seguida.
1) A replicação é a duplicação de uma molécula de DNA. Isso ocorre porque nossas células estão constantemente em divisão, e como todas as células somáticas possuem a mesma quantidade de DNA, precisamos sempre duplicar nosso DNA antes da célula se dividir.
O primeiro passo é o rompimento das ligações de hidrogênio entre as bases nitrogenadas dos nucleotídeos, permitindo a separação das duas fitas. Esse processo é auxiliado pela enzimaDNA helicase. E como a DNA helicase sabe onde ela vai começar a “abrir” o DNA? Nós possuímos em cada cromossomo uma região denominada origem de replicação, composta por uma sequência de nucleotídeos que a DNA helicase reconhece. Então, toda vez que a célula duplica seu DNA, ela começa no mesmo local.
Cada fita de DNA servirá como molde para a síntese de uma nova fita de DNA. Esse é o segundo passo, quando atuam as DNA polimerases. Essas enzimas ligam nucleotídeos que estavam dispersos no núcleo aos nucleotídeos das fitas de DNA, obedecendo às regras de pareamento entre as bases nitrogenadas. Com isso cada dupla fita de DNA nova formada será metade antiga e metade nova, e por isso que se diz que a duplicação do DNA é semiconservativa. Existem outras enzimas que atuam nesse processo, como as DNA primases que adicionam os primeiros nucleotídeos antes da DNA polimerase, além de existirem diversos tipos de DNA polimerases. O interessante é que as polimerases, além de adicionarem os nucleotídeos, possuem a capacidade de verificar se acabaram de cometer erros, a chamada atividade revisora. Mesmo com todo esse cuidado, as polimerases erram e causam muitas das nossas mutações no genoma.
A replicação sempre ocorre em um sentido: 5’- 3’, isso quer dizer que vai do grupo fosfato de um nucleotídeo (que está ligado ao carbono 5’ do açúcar) para o grupo hidroxila de outro (que está ligado ao carbono 3’ do açúcar). Com isso, a polimerase consegue sintetizar uma cadeia de maneira contínua, mas a outra não (as fitas ficam em sentidos antiparalelos). Esta fita retardada é formada aos pouquinhos, e cada fragmento que é formado é dado o nome de fragmento de Okazaki, que foi o pesquisador que os descobriu.
2) A transcrição é um processo onde a informação sai do genoma de DNA para a formação de mRNAs, que comandam toda a maquinaria celular. Como o “idioma” do DNA e do RNA é o mesmo, os nucleotídeos, a informação é transcrita, ou seja, copiada.
No caso da transcrição, a enzima que atua é aRNA polimerase, que também atua no sentido 5’-3’, mas que não precisa da enzima primase para iniciar a polimerização. Essa enzima não possui atividade revisora, mas isso não é um grande problema, pois um erro em uma molécula de RNA produzirá algumas proteínas defeituosas, ao contrário de um erro no DNA. Uma das fitas de DNA aberta serve de molde para a síntese de uma cadeira de RNA mensageiro complementar a fita molde, e que codifica pra um gene que será expresso na forma de proteína. E como a RNA polimerase sabe onde começar? A maioria dos nossos genes possuem regiões que controlam sua própria expressão, os promotores. Os promotores são sequências de nucleotídeos onde se ligam moléculas que inibem ou ativam a transcrição. Serve também, como ponto de ligação de um complexo de proteínas que auxiliam a RNA polimerase a se ligar e agir.
Um fenômeno interessante é o splicing alternativo que ocorre no processamento do mRNA ainda no núcleo. Nesse processo algumas partes não importantes para a proteína a ser formada são retiradas (íntrons), bem como é permitida a combinação entre o restante do mRNA (éxons), formando várias proteínas diferentes a partir de uma mesma molécula de mRNA.
3) A Tradução é o processo final de cascata, que ocorre nos ribossomos, livres ou no retículo endoplasmático. As moléculas de RNA são críticas nesse momento, pois são elas que fazem a ponte entre a sequência dos nucleotídeos no DNA e o “idioma” das proteínas, os aminoácidos.
Uma molécula de mRNA já processada é exportada para o citoplasma, onde se liga aos ribossomos. Lembre-se que os ribossomos, além de proteínas, são formados por moléculas de rRNA. Nos ribossomos, além do sítio para a ligação do mRNA, existem sítios para a ligação dos tRNAs, que se ligam aos nucleotídeos do mRNA. No ribossomo ocorre então a ligação entre os aminoácidos de vários tRNAs diferentes até a formação da cadeia polipeptídica, ou seja, da proteína.
Cada três letras (nucleotídeos) no DNA correspondem a uma letra (aminoácido) na proteína final, e algumas combinações diferentes de letras do DNA resultam na mesma letra da proteína, como se fossem palavras sinônimas. Por causa desse fenômeno é dito que o código genético édegenerado.
As proteínas então seguirão suas funções na célula até serem degradadas. Quando são necessárias novas enzimas, a célula novamente transcreve e traduz.
Como surgem os plastos
Os plastos surgem, basicamente, a partir de estruturas citoplasmáticas denominadas proplastos, pequenas bolsas esféricas, com cerca de 0,2 micrometros de diâmetro, delimitadas por duas membranas. No interior dos proplastos existem DNA, enzimas e ribossomos, mas não há tilacóides nem clorofila. Os proplastos são capazes de se dividir e são herdados de geração em geração celular, transmitindo-se de pais para filhos pelos gametas.
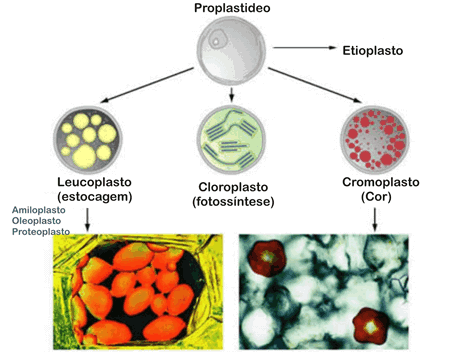
Origem dos cloroplastos
Nas células vegetais que ficam expostas à luz, como as das folhas, por exemplo, os proplastos crescem e se transformam em cloroplastos. A necessidade de luz para a sua formação explica porque não existem cloroplastos nas células das partes não iluminadas das plantas, como as das raízes ou as das partes internas dos caules. Se deixarmos uma semente germinar no escuro, as folhas da planta recém nascida serão amareladas, e em suas células não serão encontrados cloroplastos maduros, mas sim estioplasto.
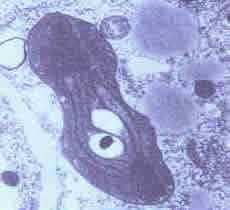
Cloroplasto e Estioplasto
Amiloplastos ou grãos de amido
Em certas situações, os cloroplastos ou os leucoplastos podem acumular grandes quantidades de amido, um polissacarídeo sintetizado a partir da glicose. O amido pode ocupar totalmente o interior da organela, que se transforma em uma estrutura conhecida como amiloplasto ou grão de amido. Os amiloplastos são grandes reservatórios de amido, que em momentos de necessidade (se faltar glicose) pode se reconvertida em glicose e utilizado.
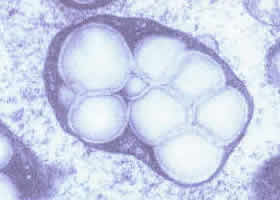
Amiloplasto
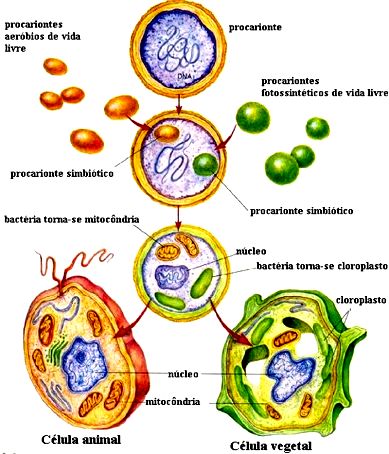
A capacidade de multiplicação dos plastos e suas semelhanças bioquímicas com os seres procariontes atuais sugerem que essas organelas tiveram como ancestrais bactérias fotossintetizantes primitivas, que há centenas de milhões de anos estabeleceram uma relação de cooperação com células eucariontes. No decorrer do processo evolutivo, a dependência entre os dois tipos de organismos teria se tornado tão grande que as bactérias fotossintetizantes e a célula eucarionte hospedeira perderam a capacidade de viver isoladamente.
Plastos
Classificação e estrutura dos plastos
Plastos são orgânulos citoplasmáticos encontrados nas células de plantas e de algas. Sua forma e tamanho variam conforme o tipo de organismo. Em algumas algas, cada célula possui um ou poucos plastos, de grande tamanho e formas características. Já em outras algas e nas plantas em geral, os plastos são menores e estão presentes em grande número por célula.
Os plastos podem ser separados em duas categorias:
- cromoplastos (do grego chromos, cor), que apresentam pigmentos em seu interior. O cromoplasto mais freqüente nas plantas é o cloroplasto, cujo principal componente é a clorofila, de cor verde. Há também plastos vermelhos, os eritroplastos(do grego eritros, vermelho), que se desenvolvem, por exemplo, em frutos maduros de tomate.
- leucoplastos (do grego leukos, branco), que não contêm pigmentos.
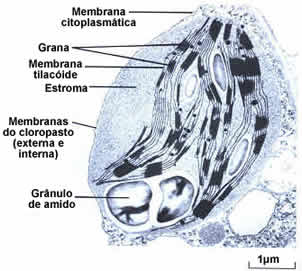
Cloroplastos
Os cloroplastos são orgânulos citoplasmáticos discóides que se assemelham a uma lente biconvexa com cerca de 10 micrometros de diâmetro. Eles apresentam duas membranas envolventes e inúmeras membranas internas, que formam pequenas bolsas discoidais e achatadas, os tilacóides (do grego thylakos, bolsa).
Os tilacóides se organizam uns sobre os outros, formando estruturas cilíndricas que lembram pilhas de moedas. Cada pilha é um granum, que significa grão, em latim (no plural, grana).
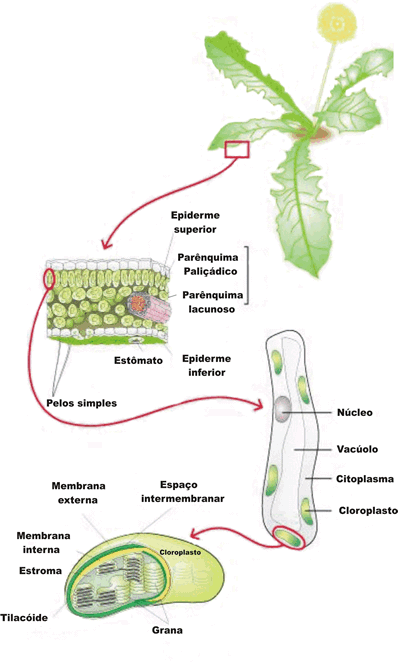
O espaço interno do cloroplasto é preenchido por um fluido viscoso denominado estroma, que corresponde à matriz das mitocôndrias, e contém, como estas, DNA, enzimas e ribossomos.
As moléculas de clorofila ficam dispostas organizadamente nas membranas dos tilacóides, de modo a captarem a luz solar com a máxima eficiência.
Funções do cloroplasto
Se as mitocôndrias são as centrais energéticas das células, os cloroplastos são as centrais energéticas da própria vida. Eles produzem moléculas orgânicas, principalmente glicose, que servem de combustível para as mitocôndrias de todos os organismos que se alimentam, direta ou indiretamente, das plantas.
Os cloroplastos produzem substâncias orgânicas através do processo de fotossíntese. Nesse processo, a energia luminosa é transformada em energia química, que fica armazenada nas moléculas das substâncias orgânicas fabricadas. As matérias-primas empregadas na produção dessas substâncias são, simplesmente, gás carbônico e água.
Durante a fotossíntese, os cloroplastos também produzem e liberamgás oxigênio (O2), necessário à respiração tanto de animais quanto de plantas. Os cientistas acreditam que praticamente todo o gás oxigênio que existe hoje na atmosfera terrestre tenha se originado através da fotossíntese.
A origem das mitocôndrias
Toda mitocôndria surge da reprodução de uma outra mitocôndria. Quando a célula vai se dividir, suas mitocôndrias se separam em dois grupos mais ou menos equivalentes, que se posicionam em cada um dos lados do citoplasma.
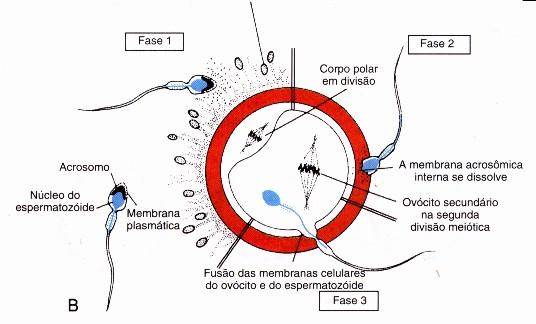
Ao final da divisão cada um dos grupos fica em uma célula-filha. Posteriormente, no decorrer do crescimento das células, as mitocôndrias se duplicam e crescem, restabelecendo o número original.
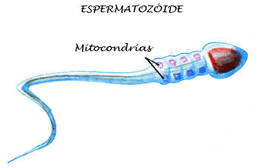 |
As mitocôndrias do espermatozóide penetram no óvulo durante a fecundação e degeneram-se logo em seguida, portanto, as mitocôndrias presentes na célula-ovo são originárias exclusivamente da mãe. As mitocôndrias ovulares, que se multiplicam sempre que a célula se reproduz, são as ancestrais de todas, as mitocôndrias presentes em nossas células.
Muitos cientistas estão convencidos de que as mitocôndrias são descendentes de seres procariontes primitivos que um dia se instalaram no citoplasma das primeiras células eucariontes. Existem evidências que apóiam essa hipótese, tais como o fato de as mitocôndrias apresentarem material genético mais parecido com a das bactérias do que com a das células eucariontes em que se encontram.
|
O mesmo ocorre com relação a maquinaria para a síntese de proteínas: os ribossomos mitocondriais são muito semelhantes aos das bactérias e bem diferentes dos ribossomos presentes no citoplasma das células eucariontes.
 ~
~
Mitocôndrias
Estrutura e função das mitocôndrias
As mitocôndrias estão imersas no citosol, entre as diversas bolsas e filamentos que preenchem o citoplasma das células eucariontes. Elas são verdadeiras “casas de força” das células, pois produzem energia para todas as atividades celulares.
As mitocôndrias foram descobertas em meados do século XIX, e, durante décadas, sua existência foi questionada por alguns citologistas. Somente em 1890 foi demonstrada, de modo incontestável, a presença de mitocôndrias no citoplasma celular. O termo “mitocôndria” (do grego, mitos, fio, e condros, cartilagem) surgiu em 1898, possivelmente como referência ao aspecto filamentoso e homogêneo (cartilaginoso) dessas organelas em alguns tipos de células, quando observadas ao microscópio óptico.
As mitocôndrias, cujo número varia de dezenas até centenas, dependendo do tipo de célula, estão presentes praticamente em todos os seres eucariontes, sejam animais, plantas, algas, fungos ou protozoários.
Estrutura interna das mitocôndrias
As mitocôndrias são delimitadas por duas membranas lipoprotéicassemelhantes às demais membranas celulares. Enquanto amembrana externa é lisa, a membrana interna possui inúmeras pregas – as cristas mitocondriais – que se projetam para o interior da organela.
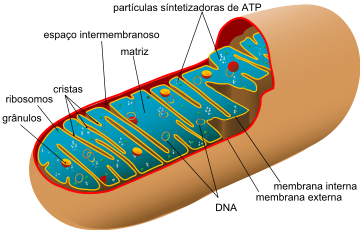
A cavidade interna das mitocôndrias é preenchida por um fluido denominado matriz mitocondrial, onde estão presentes diversas enzimas, além de DNA e RNA e pequenos ribossomos e substâncias necessárias à fabricação de determinadas proteínas.
A respiração celular
No interior das mitocôndrias ocorre a respiração celular, processo em que moléculas orgânicas de alimento reagem com gás oxigênio (O2), transformando-se em gás carbônico (CO2) e água (H2O) e liberando energia.
C6H12O6 + O2 -> 6 CO2 + 6 H2O + energia
|
A energia liberada na respiração celular é armazenada em uma substância chamada ATP (adenosina trifosfato), que se difunde para todas as regiões da célula, fornecendo energia para as mais diversas atividades celulares. O processo de respiração celular será melhor explicado na seção de Metabolismo energético.
Os centríolos
Os centríolos são organelas NÃO envolvidas por membrana e que participam do progresso de divisão celular. Nas células de fungos complexos, plantas superiores (gimnospermas e angiospermas) e nematóides não existem centríolos. Eles estão presentes na maioria das células de animais, algas e vegetais inferiores como as briófitas (musgos) e pteridófitas (samambaias).
Estruturalmente, são constituídos por um total de nove trios de microtúbulos protéicos, que se organizam em cilindro.
São autoduplicáveis no período que precede a divisão celular, migrando, logo a seguir, para os pólos opostos da célula.
Uma das providências que a fábrica celular precisa tomar é a construção de novas fábricas, isto é, a sua multiplicação. Isso envolve uma elaboração prévia de uma serie de “andaimes” protéicos, o chamado fuso de divisão, formado por inúmeros filamentos de microtúbulos.
Embora esses microtúbulos não sejam originados dos centríolos e sim de uma região da célula conhecido como centrossomo, é comum a participação deles no processo de divisão de uma célula animal. Já em células de vegetais superiores, como não existem centríolos, sua multiplicação se processa sem eles.
|  |
Os Cílios e Flagelos
São estruturas móveis, encontradas externamente em células de diversos seres vivos. Os cílios são curtos e podem ser relacionados à locomoção e a remoção de impurezas. Nas células que revestem a traquéia humana, por exemplo, os batimentos ciliares empurram impurezas provenientes do ar inspirado, trabalho facilitado pela mistura com o muco que, produzido pelas células da traquéia, lubrifica e protege a traquéia. Em alguns protozoários, por exemplo, o paramécio, os cílios são utilizados para a locomoção.
Os flagelos são longos e também se relacionam a locomoção de certas células, como a de alguns protozoários (por exemplo, o tripanosssomo causador da doença de Chagas) e a do espermatozóide.
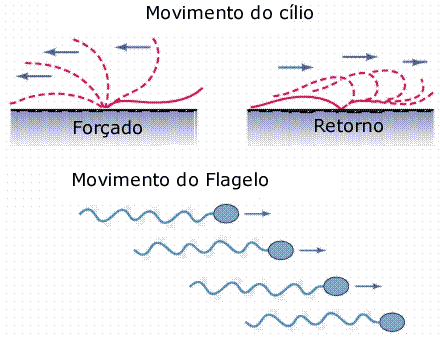
Em alguns organismos pluricelulares, por exemplo, nas esponjas, o batimento flagelar cria correntes de água que percorrem canais e cavidades internas, trazendo, por exemplo, partículas de alimento.
Estruturalmente, cílios e flagelos são idênticos. Ambos são cilíndricos, exteriores as células e cobertos por membrana plasmática. Internamente, cada cílio ou flagelo é constituído por um conjunto de nove pares de microtúbulos periféricos de tubulina, circundando um par de microtúbulos centrais. É a chamada estrutura 9 + 2.
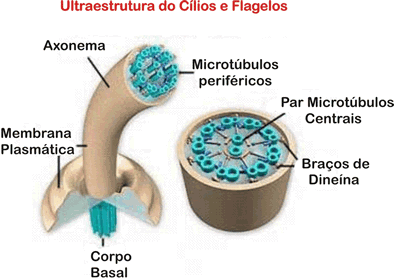
Tanto os cílios como flagelos são originados por uma região organizadora no interior da célula, conhecida como corpúsculo basal. Em cada corpúsculo basal há um conjunto de nove trios de microtúbulos (ao invés de duplas, como nos cílios e flagelos), dispostos em círculo. Nesse sentido, a estrutura do corpúsculo basal é semelhante à de um centríolo.
Filamentos intermediários
Os filamentos intermediários são assim chamados por terem um diâmetro intermediário – cerca de 10 nm – em relação aos outros dois tipos de filamentos protéicos.
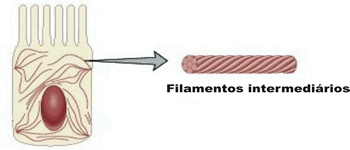
Nas células que revestem a camada mais externa da pele existe grande quantidade de um tipo de filamento intermediário chamadoqueratina. Um dos papeis desse filamento é impedir que as células desse tecido se separem ou rompam ao serem submetidas, por exemplo, a um estiramento.
Além de estarem espalhadas pelo interior das células, armando-as, moléculas de queratina promovem uma “amarração” entre elas em determinados pontos, o que garante a estabilidade do tecido no caso da ação de algum agente externo que tente separá-las. Esse papel é parecido ao das barras de ferro que são utilizadas na construção de uma coluna de concreto. Outras células possuem apreciável quantidade de outros filamentos intermediários. É o caso das componentes dos tecidos conjuntivos e dos neurofilamentos encontrados no interior das células nervosas.
Resumo
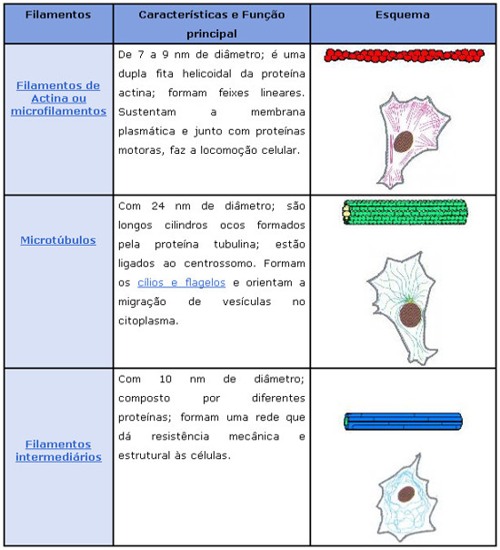
Citoesqueleto
Quando se diz que o hialoplasma é um fluido viscoso, fica-se com a impressão de que a célula animal tem uma consistência amolecida e que se deforma a todo o momento. Não é assim.
Um verdadeiro “esqueleto” formado por vários tipos de fibras de proteínas cruza a célula em diversas direções, dando-lhe consistência e firmeza.
Essa “armação” é importante se lembrarmos que a célula animal é desprovida de uma membrana rígida, como acontece com a membrana celulósica dos vegetais.
Entre as fibras protéicas componentes desse “citoesqueleto” podem ser citados os microfilamentos de actina, os microtúbulos e osfilamentos intermediários.
Os microfilamentos são os mais abundantes, constituídos daproteína contráctil actina e encontrados em todas as células eucarióticas. São extremamente finos e flexíveis, chegando a ter 3 a 6 nm (nanômetros) de diâmetro, cruzando a célula em diferentes direções , embora concentram-se em maior número na periferia, logo abaixo da membrana plasmática. Muitos movimentos executados por células animais e vegetais são possíveis graças aos microfilamentos de actina.
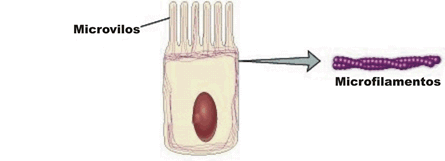
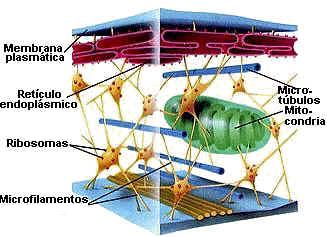
Os microtúbulos, por sua vez, são filamentos mais grossos, de cerca de 20 a 25 nm de diâmetro, que funcionam como verdadeiros andaimes de todas as células eucarióticas. São, como o nome diz, tubulares, rígidos e constituídos por moléculas de proteínas conhecidas como tubulinas, dispostas helicoidalmente, formando um cilindro. Um exemplo, desse tipo de filamento é o que organiza o chamado fuso de divisão celular. Nesse caso, inúmeros microtúbulos se originam e irradiam a partir de uma região da célula conhecida como centrossomo (ou centro celular) e desempenham papel extremamente importante na movimentação dos cromossomos durante a divisão de uma célula.
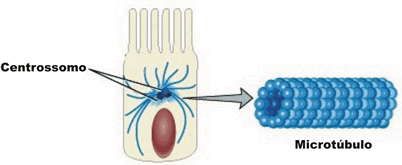
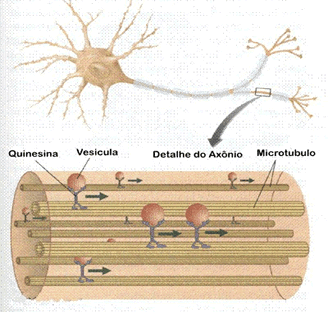
Outro papel atribuído aos microtúbulos é o de servir como verdadeiras “esteiras” rolantes que permitem o deslocamento de substâncias, de vesículas e de organóides como as mitocôndrias e cloroplastos pelo interior da célula. Isso é possível a partir da associação de proteínas motoras com os microtúbulos.
Essas proteínas motoras ligam-se de um lado, aos microtúbulos e, do outro, à substância ou organóide que será transportado, promovendo o seu deslocamento.
Por exemplo, ao longo do axônio (prolongamento) de um neurônio, as proteínas motoras conduzem, ao longo da “esteira” formada pelos microtúbulos, diversas substâncias para as terminações do axônio e que terão importante participação no funcionamento da célula nervosa.
|  |
Peroxissomos
Peroxissomos são bolsas membranosas que contêm alguns tipos de enzimas digestivas. Sua semelhança com os lisossomos fez com que fossem confundidos com eles até bem pouco tempo. Entretanto, hoje se sabe que os peroxissomos diferem dos lisossomos principalmente quanto ao tipo de enzimas que possuem.
Os peroxissomos, além de conterem enzimas que degradam gorduras e aminoácidos, têm também grandes quantidades da enzima catalase.
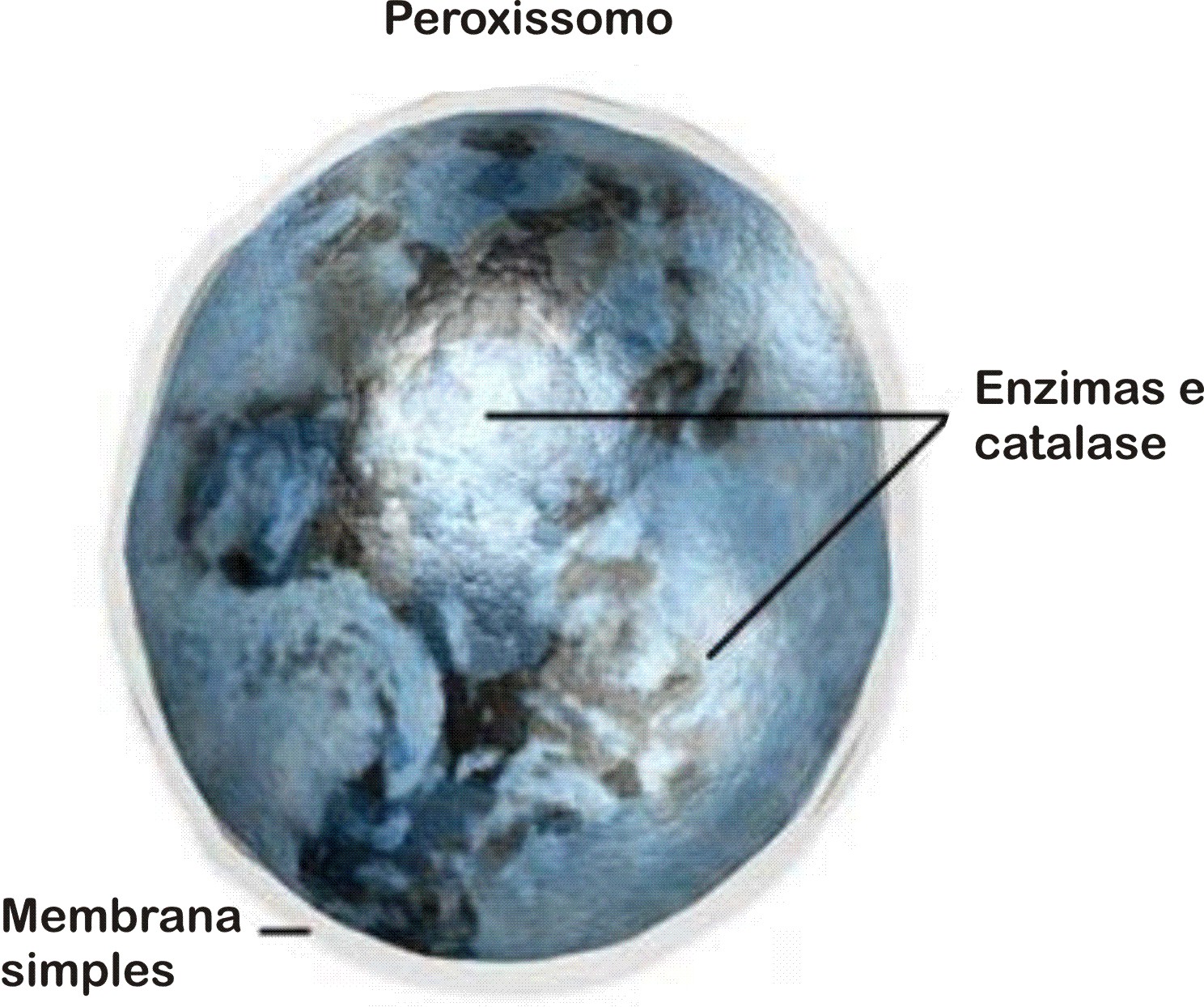
A catalase converte o peróxido de hidrogênio, popularmente conhecido como água oxigenada (H2O2), e água e gás oxigênio. A água oxigenada se forma normalmente durante a degradação de gorduras e de aminoácidos, mas, em grande quantidade, pode causar lesões à célula.
2 H2O2 + Enzima Catalase → 2 H2O + O2
|
Apesar das descobertas recentes envolvendo os peroxissomos, a função dessas organelas no metabolismo celular ainda é pouco conhecida. Entre outras funções, acredita-se que participem dos processos de desintoxicação da célula.
Glioxissomos
Em vegetais, as células das folhas e das sementes em germinação possuem peroxissomos especiais, conhecidos como glioxissomos. Nas células das folhas, essas estruturas atuam em algumas reações do processo de fotossíntese, relacionadas à fixação do gás carbônico. Nas sementes, essas organelas são importantes natransformação de ácidos graxos em substâncias de menor tamanho, que acabarão sendo convertidas em glicose e utilizadas pelo embrião em germinação.
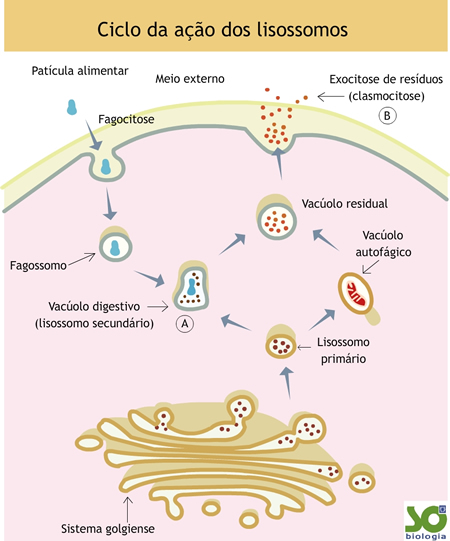
Vacúolos digestivos
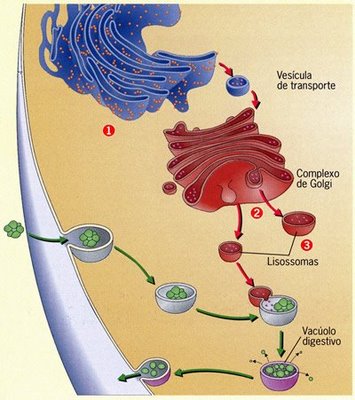 |
As bolsas originadas pela fusão de lisossomos com fagossomos ou pinossomos são denominadasvacúolos digestivos; em seu interior, as substâncias originalmente presentes nos fagossomos ou pinossomos são digeridas pelas enzimas lisossômicas.
À medida que a digestão intracelular vai ocorrendo, as partículas capturadas pelas células são quebradas em pequenas moléculas que atravessam a membrana do vacúolo digestivo, passando para o citosol. Essas moléculas serão utilizadas na fabricação de novas substâncias e no fornecimento de energia à célula.
Eventuais restos do processo digestivo, constituídos por material que não foi digerido, permanecem dentro do vacúolo, que passa a ser chamado vacúolo residual.
Muitas célula eliminam o conteúdo do vacúolo residual para o meio exterior. Nesse processo, denominado clasmocitose, o vacúolo residual encosta na membrana plasmática e fundem-se com ela, lançando seu conteúdo para o meio externo.
|
Autofagia
Todas as células praticam autofagia (do grego autos, próprio, ephagein, comer), digerindo partes de si mesmas com o auxílio de seus lisossomos. Por incrível que pareça, a autofagia é uma atividade indispensável à sobrevivência da célula.
Em determinadas situações, a autofagia é uma atividade puramente alimentar. Quando um organismo é privado de alimento e as reservas do seu corpo se esgotam, as células, como estratégia de sobrevivência no momento de crise, passam a digerir partes de si mesmas.
No dia-a-dia da vida de uma célula, a autofagia permite destruir organelas celulares desgastadas e reaproveitar alguns de seus componentes moleculares.
O processo da autofagia se inicia com a aproximação dos lisossomos da estrutura a ser eliminada. Esta é cercada e envolvida pelos lisossomos, ficando contida em uma bolsa repleta de enzimas denominada vacúolo autofágico.
Através da autofagia, uma célula destrói e reconstrói seus constituintes centenas ou até milhares de vezes. Uma célula nervosa do cérebro, por exemplo, formada em nossa vida embrionária, tem todos os seus componentes (exceto os genes) com menos de um mês de idade. Uma célula de nosso fígado, a cada semana, digere e reconstrói a maioria de seus componentes.
Na silicose (“doença dos mineiros”), que ataca os pulmões ocorre a ruptura dos lisossomos de células fagocitárias (macrófagos), com conseqüente digestão dos componentes e morte celular.
Certas doenças degenerativas do organismo humano são creditadas a liberação de enzimas lisossômicas dentro da célula; isso aconteceria, por exemplo, em certos casos de artrite, doença das articulações ósseas.
Lisossomos
Estrutura e origem dos lisossomos
Os lisossomos (do grego lise, quebra, destruição) são bolsas membranosas que contêm enzimas capazes de digerir substâncias orgânicas. Com origem no aparelho de Golgi, os lisossomos estão presentes em praticamente todas as células eucariontes. Asenzimas são produzidas no RER e migram para os dictiossomos, sendo identificadas e enviadas para uma região especial do aparelho de Golgi, onde são empacotadas e liberadas na forma de pequenas bolsas.
A digestão intracelular
Os lisossomos são organelas responsáveis pela digestão intracelular. As bolsas formadas na fagocitose e na pinocitose, que contêm partículas capturadas no meio externo, fundem-se aos lisossomos, dando origem a bolsas maiores, onde a digestão ocorrerá.
Assinar:
Comentários (Atom)